起源
共生細菌によって産生されるセルラーゼは反芻動物に見られ、最近では緑藻にも見られます.
微生物セルラーゼは、通常 Aspergillus niger を使用して、真菌の発酵から取得できます。 およびTrichoderma sp .
機能
セルラーゼは、パンの処方へのふすま添加の悪影響を最小限に抑えるのに役立ちます.ふすまは非常に価値のある栄養成分であるにもかかわらず、フォーム構造を大きく乱し、グルテンネットワークの形成を制限するため、製パン業者にさらなる技術的課題を課しています.
パンにセルラーゼを使用すると、次の属性が得られます:
- 自由水の放出によるパンの流れの増加
- 最適なオーブン スプリングと製品容量のための優れたガス保持力
- 気泡の合体を最小限に抑えるための、より強力で継続的なグルテン ネットワーク
- クラム内のガスセル構造がより細かく均一
- グルコースオキシダーゼとの強力な相乗効果(この場合、セルロースはグルコース基質を酸化酵素に提供して強化機能を強化します)
- さまざまな種類のパンの柔らかいパン粉とより長い貯蔵寿命
- 低糖繊維パンの皮の色が濃く均一
セルラーゼには主に 4 つのタイプがあります:
<オール>次の表は、セルラーゼの各タイプの重要な側面をまとめたものです:
| EC番号 | 反応触媒 | 追加コメント |
|---|---|---|
| 3.2.1.4 エンドスプリッティング酵素エンドグルカナーゼ | セルロースのβ-(1,4)-D-およびβ-(1,3)-D-グルコシド結合のランダムエンド加水分解 (グルカン) 高分子鎖。 製品には、さまざまなサイズのグルコース、セロビオース、セロデキストリンが含まれます。 | 綿などの結晶セルロースに対して不活性です。 非晶質セルロース (結晶セルロースの非晶質領域を含む) と、増粘剤カルボキシメチルセルロース (CMC) やヒドロキシメチルプロピルセルロース (HPMC) などの可溶性基質を加水分解できます。 エンドグルカナーゼ活性により、粘度が急速に低下し、メイラード反応に参加できる還元基が急激に増加します。 その作用は真菌のα-アミラーゼに非常に似ています. |
| 3.2.1.91 エキソ分裂酵素 | 1,4-β-D-グルカン セロビオヒドロラーゼ グルカン基質の非還元末端からセロビオースを連続的に除去することにより、非晶質セルロースを分解します。 | 純粋な場合、微結晶性セルロースを分解する可能性があります。 粘度の低下に対する還元基の増加率は、エンドグルカナーゼよりもはるかに高くなっています。エンドグルカナーゼとセロビオヒドロラーゼは、結晶セルロースの加水分解において相乗的に作用します。 その作用は、穀物のベータアミラーゼに非常に似ています。 |
| EC 3.2.1.74 エキソグルコヒドロラーゼ | セロデキストリンの非還元末端からグルコース単位を連続的に加水分解します。 | グルカン分子の鎖長が短くなると、加水分解の速度が低下します。 その作用はアミログルコシダーゼ (AMG) によく似ています。 |
| 3.2.1.21 β-グルコシダーゼ β-D-グルコシドグルコヒドロラーゼ | セロビオースを 2 つのグルコース単位に切断し、小さなセロデキストリンの非還元末端からグルコースを除去します。 | エキソグルコヒドロラーゼとは異なり、β-グルコシダーゼの速度はグルカン鎖のサイズが小さくなるにつれて増加し、セロビオースが最も速く加水分解されます. その作用は、マルターゼと AMG の組み合わせに非常に似ています。 |
商業生産
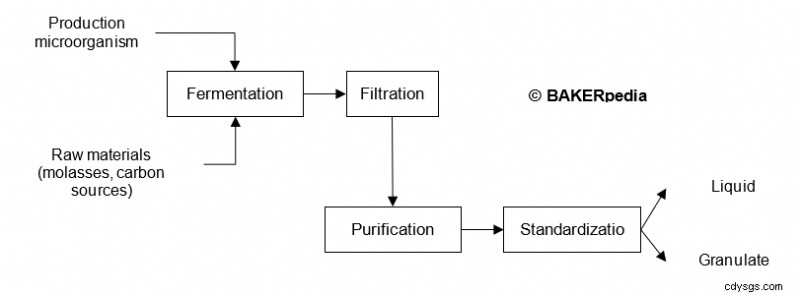
セルラーゼの大規模生産は、最新のバイオテクノロジーを使用した発酵と下流の処理に依存しています。
次のブロック図は、セルラーゼの商業生産をまとめたものです:
申し込み
菌類のセルラーゼは、分子量が約 63,000 Da、至適 pH が 5.5 ~ 6.0 で、100°C、pH 7 で 5 分間安定です。この酵素は、重金属イオン、スルフヒドリル試薬、酸化および還元によって阻害されます。
セルラーゼ活性は、セルロースから放出される還元基の速度を測定することによって決定できます。
規制
他の酵素と同様に、食品セルラーゼは GRAS (Generally Recognized as Safe) であり、米国では食品加工助剤と見なされることがよくあります。 FDA はその起源 (食品適合性) を規制し、GMP に基づいて (該当する場合) 使用制限を設定します。